シナプスは...シナプスの構造。神経系、筋肉系、化学系のシナプス
シナプスはプロセスの特定の接触領域です情報細胞の伝達を提供する神経細胞および他の非興奮性および興奮性細胞。シナプスは、2つの細胞の接触膜によって形態学的に形成される。膜は、神経細胞のプロセスに関連して、シグナルが入る細胞のシナプス前膜と呼ばれ、第二の名前はシナプス後シナプスである。シナプス後膜の付属物と共に、シナプスは、神経細胞内、神経筋肉および神経分泌性であり得る。シナプスという言葉は、Charles Sherrington(英語の生理学者)によって1897年に導入されました。

シナプスとは何ですか?
シナプスは、神経インパルスの神経線維から他の神経線維または神経細胞への移行を確実にし、レセプター細胞(神経細胞および他の神経線維との接触領域)からの神経線維には2つの神経細胞が必要である。
シナプスは、ニューロンの終わりにある小さな部門です。 彼の助けを借りて、情報は最初のニューロンから2番目のニューロンに転送されます。シナプスは、神経細胞の3つのセクションに位置しています。また、シナプスは、神経細胞が体の様々な腺または筋肉と接触する場所にある。
シナプスの構成
シナプスの構造は簡単な回路を持っています。 それは3つの部分から構成され、各部分は情報の転送中に特定の機能を実行します。したがって、このようなシナプスの構造は、神経インパルスの伝達に適していると言える。情報転送のプロセスに直接的に、2つの主要なセルが作用する:知覚者と伝達。伝達細胞の軸索の終わりには、シナプス前の終結(シナプスの初期部分)がある。神経伝達物質(この言葉はいくつかの意味を持っています:メディエーター、メディエーターまたは神経伝達物質)を誘発する細胞に影響する可能性があります - 電気信号が2つのニューロン間で伝達される特定の化学物質。

シナプスのメディエーター
メディエーター(ラテンメディアから - トランスミッター、ミドルマンまたはミドル)そのようなシナプスの媒介物質は、神経インパルスの伝達過程において非常に重要である。
阻害剤との間の形態的差異エキサイティングなシナプスは、メディエイターの解放メカニズムを持たないということです。阻害性シナプス、運動ニューロンおよび他の阻害性シナプスにおけるメディエータは、アミノ酸グリシンであると考えられる。しかし、シナプスの抑制的または興奮的な特徴は、それらのメディエーターによってではなく、シナプス後膜の特性によって決定される。例えば、アセチルコリンは、末端(神経心筋の迷走神経)の神経筋シナプスに刺激効果を与える。
アセチルコリンは、興奮性メディエーターとして作用するコリン作動性シナプス前終末にレンショウ細胞上のシナプスにおけるシナプス(シナプス前膜は、脊髄運動ニューロンの端を果たしている)、汗腺、交感神経系の神経節における腸およびシナプスにおける髄質nadpochenikov。 Atsetilholi-nesterazuおよびアセチルコリンはまた、時には大量に、脳の異なる部分の画分に見出さが、レンショウがまだ残っているコリン作動性シナプスを同定することができなかった細胞上のコリン作動性シナプス以外。科学者によると、中枢神経系の興奮伝達物質アセチルコリンの機能は非常に可能性があります。

シナプスのメディエーターが機能を完了すると、カテコールアミンはシナプス前神経末端に吸収され、一方膜貫通輸送が関与する。メディエーターの吸収の間、シナプスは、長期かつ律動的な仕事の間に予備の早期枯渇から保護される。
シナプス:主なタイプと機能
1892年のラングレーは、哺乳類の栄養神経節におけるシナプス伝達は電気的ではなく化学的である。 10年後、Eliottは、副腎からのアドレナリンが、交感神経の刺激と同じ効果から得られることを見出した。

シナプス化学
化学シナプスは根本的に異なるシナプス後のメディエイターの助けを借りて刺激を移してポストシナップにする。したがって、化学的シナプスの形態において相違が生じる。化学的シナプスは、椎骨CNSにおいてより一般的である。現在、ニューロンは一対のメディエーター(共存するメディエーター)を分泌し、合成することができることが知られている。ニューロンはまた、神経伝達物質の可塑性を有している - 発達中に主要メディエーターを変更する能力。

神経筋シナプス
このシナプスは興奮の伝達を行い、しかしながら、この関係は様々な要因を破壊する可能性がある。伝達は、アセチルコリンのシナプス間隙への放出の遮断中、またシナプス後膜のゾーンにおけるその含有量の過剰の間に終了する。多くの毒物や薬物が捕獲に影響を与え、シナプス後膜のホール受容体に関連した出力が得られ、筋シナプスは興奮伝達を遮断する。呼吸筋の収縮を止めて止める間に体が死ぬ。

シナプス神経:機能とコンポーネント
シナプスは、2人の間の接触場所の接続です細胞。それらの各々は、そのエレクトロジェニック膜に封入されている。神経シナプスは、シナプス後膜、シナプス間隙およびシナプス前膜の3つの主要成分からなる。シナプス後膜(post-synaptic membrane) - これは神経末端であり、筋肉に到達し、筋肉組織の内部に降下する。シナプス前領域には、小胞があります。これらは小腔であり、メディエーターを持っています。彼らは常に動いている。
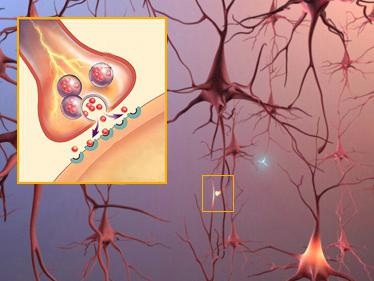
シナプス後膜とシナプス前膜の間のシナプス間隙
異なるシナプスでは、隙間の大きさが異なる。 この空間には、メディエーターが存在する細胞間液が満たされています。シナプス後膜は、神経終末が筋神経シナプス中の神経支配細胞と接触する場所をカバーする。ある種のシナプスではシナプス後膜がしわを生じ、接触面積が増加する。
シナプス後膜を構成する追加の物質
シナプス後膜のゾーンには、以下の物質が存在する:
- 受容体(筋骨格シナプスにおけるホールレセプター)。
- リポタンパク質(アセチルコリン)。このタンパク質は、求電子末端およびイオンヘッドを有する。頭部はシナプス間隙に入り、アセチルコリンのカチオン性頭部との相互作用が起こる。この相互作用のために、シナプス後膜に変化があり、脱分極が起こり、潜在的に依存するNaチャネルが明らかになる。膜の脱分極は自己補強プロセスとはみなされない。
- 徐々に、シナプス後膜上のその潜在能力は、メディエーターの数に依存する。すなわち、潜在的なものは局所的興奮の性質によって特徴付けられる。
- コリンエステラーゼ - はタンパク質とみなされ、酵素機能があります。構造上、コリン作動性受容体に類似しており、アセチルコリンと同様の性質を有する。コリンエステラーゼは、コリン作動性受容体に関連するアセチルコリンを最初に破壊した。コリンエステラーゼの作用下で、ホリンレセプターがアセチルコリンを除去すると、シナプス後膜の再分極が形成される。アセチルコリンは、筋肉組織の栄養に必要な酢酸およびコリンに分解される。
現在の輸送手段の助けを借りてシナプス前膜コリンは、新しいメディエーターを合成するために使用されます。メディエーターの影響下で、シナプス後膜の透過性が変化し、コリンエステラーゼ感受性および透過性の下で初期値に戻る。化学受容体は新しいメディエーターと相互作用することができる。
</ p>